
Experimental models of osteoarthritis
Last reviewed: 23.04.2024

All iLive content is medically reviewed or fact checked to ensure as much factual accuracy as possible.
We have strict sourcing guidelines and only link to reputable media sites, academic research institutions and, whenever possible, medically peer reviewed studies. Note that the numbers in parentheses ([1], [2], etc.) are clickable links to these studies.
If you feel that any of our content is inaccurate, out-of-date, or otherwise questionable, please select it and press Ctrl + Enter.
Cartilage is a highly specialized tissue containing only one type of cells (chondrocytes), characterized by the absence of blood and lymphatic vessels. Nutrition of the cartilage is mainly carried out by absorption from the synovial fluid. Metabolism of chondro-cytes is regulated by a number of soluble factors produced locally by chondrocytes and surrounding tissues. The function of chondrocytes also depends on the composition of the extracellular medium (oxygen tension, ion concentration, pH, etc.), VCM composition, cell and matrix interaction, physical signals. The main task of experimental modeling is the creation of cultures in the extracellular environment without changing the phenotype of mature cells. The second task is to create cultures for studying the premature, delayed, short or long-term response of chondrocytes to chemical and / or physical signals. Studies in vitro also provide an opportunity to study the behavior of chondrocyte in osteoarthritis. The third task is the development of co-curative systems, which allow studying the interactions of various tissues in the joint. The fourth task is the preparation of cartilaginous implants for the subsequent transplantation. And, finally, the fifth task is to study growth factors, cytokines or therapeutic agents that are capable of stimulating repair and / or inhibiting its resorption of cartilage.
Over the past decades, various models of articular cartilage cell cultures have been created, including monolayer cultures, suspended cultures, chondron cultures, explants, cocultures, immortal cell cultures. Each culture has its advantages and disadvantages and each is suitable for studying one particular aspect of chondrocyte metabolism. Thus, cartilaginous explants are an excellent model for studying the turnover of matrix elements, which requires genuine cell surface receptors and normal cell-matrix and matrix-cell interactions. At the same time, the study of deposits in the matrix or mechanisms for the regulation of chondrocyte metabolism is recommended to be carried out on a culture of isolated cells. A monolayer low-density culture is necessary for studying the process of cell differentiation. Cultures suspended in a natural or synthetic matrix are a model for analyzing the adaptive response of chondrocytes to mechanical stress.
Chondrocyte cultures
When choosing cartilage tissue for in vitro studies, several important points need to be considered. Matrix composition and metabolic activity of chondrocytes vary in different joints, and the latter also depends on the depth of the chondrocyte in the tissue. These data were obtained in several experiments in which isolated subpopulations of chondrocytes from the cartilage zones of different depths were studied. A number of morphological and biochemical differences were found between cultivated chondrocytes located in the surface and deep layers of the articular cartilage. Surface cells synthesize a rare, depleted proteoglycan fibrillar matrix, while deeper cells produce a matrix that is rich in fibrils and proteoglycans. Moreover, the surface cells produce relatively more small non-aggregated proteoglycans and hyaluronic acid and relatively less aggrecan and keratan sulfate than the more deeply located chondrocytes. Another important distinguishing feature of the metabolism of chondrocytes isolated from the cartilage zones of different depths is the response to the exogenous stimulus. According to M. Aydelotte and co-authors, the bull chondrocytes from the surface zone of the cartilage were more sensitive to IL-1 than the cells of the deep zone.
The behavior of the cells also depends on the location of the tissue. The chondrocytes of the cartilages of the ribs and ears, taken from the same animal, respond differently to growth factors, such as fibroblast growth factor (FGF) and TGF-beta. FGF increased the incorporation of thymidine, proline and leucine into the chondrocyte culture of the rib, but not the ear. TGF-P increased the incorporation of thymidine into the chondrocytes of the cartilage of the rib and ear, but did not affect the incorporation of thymidine and proline into the ear chondrocytes. Cartilage cells obtained from zones bearing the greatest load differ from those from sites with a low load on the cartilage. Thus, the chondrocytes of the mature cartilage of the knee joint of the sheep from the central region of the articular surface of the lumbar bone not covered by the meniscus, which carries the greatest load in vivo, less synthesize aggrecan, but more decors than cells from the meniscus-covered areas. The authors also emphasize the importance of using cartilage from identical joint zones when examining the synthetic function of the joints.
The metabolism of chondrocytes and their response to regulatory factors also significantly depends on the age of the donor, the development of its skeleton and the state of the joints from which the cells are taken. In human chondrocytes, a significant decrease with age of the proliferative response is observed. The greatest decrease is observed in donors aged 40-50 years and over 60 years. Moreover, the severity of the proliferative response to growth factors (eg, FGF and TGF-beta) decreases during aging. In addition to quantitative changes in the proliferation of chondrocytes, there are also qualitative changes. Young donor cells (10-20 years old) respond better to platelet derived growth factor (PDGF) than to TGF-beta, whereas the opposite is observed in adult donor cells. To explain the age-dependent changes in the synthetic function of chondrocytes and their response to the effect of growth factors, several mechanisms are used. Among them, a decrease in the number and affinity of surface cellular receptors, a change in the synthesis and bioactivity of growth factors and cytokines, a modification of postreceptor signals.
The pathological condition of the joints also alters the morphology and metabolic activity of chondrocytes. So, J. Kouri and co-authors (1996) identified three subpopulations of chondrocytes in cartilage with osteoarthritis. Chondrocytes from the superficial and upper middle of the cartilage form clusters and synthesize more proteoglycans and collagen. TGF-beta and insulin-like growth factor (IGF) are able to stimulate the synthesis of proteoglycans by chondrocytes and partially neutralize the effects of IL-1 and TNF-a. Cartilage explants affected by osteoarthritis and chondrocytes isolated from the cartilage of a patient with osteoarthritis are more sensitive to stimulation of TGF-beta than chondrocytes of healthy cartilage. These differences are most likely associated with phenotypic changes in chondrocytes in the upper layers of the articular cartilage.
Isolation of individual chondrocytes is achieved by sequential treatment with proteolytic enzymes of ECM. After their release from the ECM, isolated cells are ideally suited for studying the synthesis of de novo matrix components . Some authors use only clostridium collagenase, others pre-incubate cartilage with trypsin, pronase, DNase and / or hyaluronidase. The number of isolated cells depends on the enzymes used. Thus, when processing one of 1 g collagenase tissue can be obtained 1,4T0 6 chondrocytes, whereas when using pronase, hyaluronidase and collagenase - 4,3-10 6. When processing with collagenase, aggrecan, proteins, IL-6, IL-8 remain in the cell culture much more than in the case of sequential treatment with various enzymes. There are several explanations for these differences between the two cell cultures:
- Cell receptors are damaged or inhibited by enzymes, TGF-beta inhibits DNA and the synthesis of proteoglycans in newly isolated chondrocytes (day 1), whereas DNA and the synthesis of proteoglycans of chondrocytes cultured in a monolayer (7 days) are stimulated by TGF-beta. However, to reexpress these membrane components, an adequate period is required before the start of the experiment.
- Exogenous proteases can break the interaction of cells and the matrix, mediated by integrins. The integrin family promotes the attachment of chondrocytes to VKM molecules (Shakibaei M. Et al., 1997). This rupture can affect the expression of matrix genes.
- Residues of matrix components can regulate the synthetic function of chondrocytes. Integrins are able to recognize degradation products of ECM, thereby playing an important role in tissue repair after exposure to proteolytic enzymes. T. Larsson and co-authors (1989) reported that the addition of intact or fragmented pro-theoglycans to cell culture stimulates the synthesis of proteins and proteoglycans. However, a high level of hyaluronic acid causes a significant reduction in the inclusion of sulfates in the synthesis of proteoglycans by the chondrocytes of the chick embryo, mature chondrocytes of the pig, and rat chondrosarcoma cells. Moreover, hyaluronic acid is an inhibitor of the release of proteoglycans from cells even in the presence of IL-lb, TNF-a, FGF, which indicates a resistance to the first biological activity of growth factors and cytokines. The exact mechanism underlying the action of hyaluronic acid remains unclear; It is known that chondrocytes contain a receptor for hyaluronic acid, associated with actin filaments of the cytosol. The binding of hyaluronic acid to its receptor stimulates the phosphorylation of proteins. Thus, these data demonstrate the modulation of the metabolic function of chondrocytes by fragmented or native molecules of matrix proteins by activating the membrane receptor cells.
- The rapid stimulation by enzymes of the synthesis of matrix proteins by chondrocytes can be a consequence of a change in the shape of chondrocytes and / or the reorganization of the cytoskeleton.
- Some cytokines (eg, IL-8) and growth factors (eg, IGF-1, TGF-P) are fixed in the ECM. The best known example is the binding of TGF-beta with decore, which leads to a decrease in the ability of the former to induce cellular growth in ovarian cells in Chinese hamsters. The data that the content of cartilage decoration increases with age, indicate a decrease in bioavailability of TGF-beta in aging. Growth factors and cytokines can be released from matrix residues during culture and then modulate chondrocyte function.
Monolayer culture of chondrocytes
The differentiated phenotype of chondrocytes is primarily characterized by the synthesis of type II collagen and tissue-specific proteoglycans, as well as a low level of mitotic activity. There is evidence that with prolonged cultivation of cells in a monolayer, and after several repeated passages of cells, the chondrocytes lose their spherical outlines, acquire an elongated, fibroblast-like shape. With this fibroblast metaplasia, the synthetic function of cells is also modified, characterized by a progressive decrease in the synthesis of collagen II, IX, and XI types and an increase in the synthesis of collagens I, III and Utyopov. Small non-aggregated proteoglycans are synthesized by functional aggrecan. Synthetzatepsin B and L is extremely low in differentiated cells, but in the process of loss of differentiation increases. Collagenase-1 is expressed in differentiated chondrocytes, with prolonged cultivation, its expression decreases, while the production of tissue inhibitors of metalloproteases (TIMP) increases.
The differentiated chondrocytes re-express the collagen of the differentiated phenotype when they are transferred from a monolayer culture to a suspended one. The process of differentiation is probably related to the shape of the cells. This property is regularly used by researchers who study defective transplants with autologous chondrocytes. A small number of cells obtained from a biopsy material can be multiplied in a monolayer culture and then placed again in a three-dimensional matrix before transplantation. Re-expression of a specific phenotype by dedifferentiated chondrocytes transferred to an agarose culture can be stimulated with TGF-p, an ossein-hydroxyapatite complex and ascorbic acid.
In response to the effect of growth factors and cytokines, chondrocytes are modified during the differentiation process. The cellular response to cytokines and growth factors differ between undifferentiated and differentiated chondrocytes. IL-1 stimulates the proliferation of fibroblasts, while the growth of undifferentiated chondrocytes is inhibited by IL-1. Synthesis of DNA is stimulated by IGF-1 in elongated, but not flattened chondrocytes. In the differentiated chondrocytes, the stimulating effects of IL-1β and TNF-α on procollagenase products are more pronounced than in undifferentiated ones.
Cultivation of chondrocytes
The cultivation of chondrocytes in suspension in a liquid medium or in a natural or synthetic three-dimensional matrix stabilizes the phenotype of the chondrocyte. Cells retain their spherical shape, synthesize tissue-specific proteins. A weighted chondrocyte culture is usually recommended for the study of the formation of a new pericellular matrix. Chondrocyte cultures in synthetic or natural absorbent polymers are used to implant cells into cartilage defects to stimulate the regeneration of the cartilage tissue of the joint. Synthetic or natural environment for implantable cells must satisfy a number of requirements:
- Implants should have a porous structure for adhesion and cell growth,
- neither the polymer itself nor the products of its degradation should cause inflammation or toxic reactions during in vivo implantation ,
- the transplant carrier should be able to bind to an adjacent cartilage or subchondral bone,
- a natural or synthetic matrix must be capable of absorption, its degradation must be balanced by tissue regeneration,
- To facilitate cartilage repair, the chemical structure and matrix architecture of the matrix should help maintain the cellular phenotype inserted in the chondrocytes and the synthesis of tissue-specific proteins,
- during implantation in vivo, it is necessary to study the mechanical properties of the synthetic or natural matrix.
Suspension of chondrocytes in the liquid phase
Attaching cells to plastic vessels in which chondrocytes are cultivated can be prevented by coating their walls with a solution of methylcellulose, agarose, hydrogel (poly-2-hydroxyethyl methacrylate) or a mixture of collagen-agarose. Under these conditions, chondrocytes form clusters and synthesize mainly aggrecan and tissue-specific collagens (II, IX, XI types). Usually, two types of cells are found. The cells located in the center retain a spherical shape surrounded by a well developed ECM, which is confirmed by histochemical and ultrastructural studies. On the periphery chondrocytes have discoid contours, are surrounded by a rare ECM; Little is known about the functional characteristics of such cells.
Cultivation of chondrocytes on microcarriers supported in suspension is possible; Dextran beads (cytodex), collagen-coated dextran beads (cytodex III), non-void microspheres of collagen type I (collagen) are used as microcarriers. Under these culture conditions, chondrocytes attach to the surface of the microcarrier, retain their spherical shape, and produce a matrix-like material. Moreover, the use of collagen promotes the proliferation of chondrocytes and the reexpression of a normal phenotype. Therefore, the cultivation of chondrocytes on the microspheres of the collagen can be used to restore the cell phenotype before transplantation.
Another method of cultivation of a suspension of chondrocytes in a liquid medium is their cultivation in the form of dense beads consisting of cells (0.5-1 * 10 b ) obtained by centrifugation. Such chondrocytes are able to produce a matrix containing a large number of proteoglycans, collagen type II, but not type I collagen, which is confirmed by histological, immunohistochemical and quantitative methods.
Suspension of chondrocytes in a natural ECM
Chondrocytes can be cultured in suspension in a three-dimensional matrix (soft agar, agarose, collagen gel or sponge, hyaluronic acid, fibrin glue, alginate beads).
Cultured agarose chondrocytes retain their normal phenotype and synthesize collagen type II and tissue-specific aggregate-new aggregates. When cultured in agarose, cell-synthesized proteoglycans are released into the medium for 50 days. For comparison - in monolayer culture the cell phase is overfilled with glycosaminoglycans already in the first 5-6 days of cultivation; when cultivated in the medium after the synthesis and release of glycosaminoglycans is intensified, the time-dependent decrease in glycosaminoglycans occurs in the first 8-10 days. Nevertheless, the behavior of chondrocytes during their cultivation in agarose differs from that in in vivo conditions. In agarose, a large number of synthesized Aggregan aggregates contain smaller and smaller molecules than in vivo. TGF-P stimulates the synthesis of proteoglycans in the explant, but reduces the synthesis of aggrecan in agarose.
Alginate is a linear polysaccharide derived from brown seaweed. In the presence of divalent cations, such as Ca 2+ ions, this polymer becomes a gel. Each chondrocyte found in the alginate is surrounded by a matrix of negatively charged polysaccharides whose pores are commensurate with those in the hyaline cartilage. The matrix that forms the chondrocyte in the alginate beads consists of two sections - a thin layer of the cell-associated matrix corresponding to the pericellular and territorial matrix, articular cartilage and a more remote matrix equivalent to the interterritorial matrix in the native tissue. On the 30th day of cultivation, the relative and absolute volume occupied by the cells, and each of the two divisions in the alginate bead are almost identical to those in the native cartilage. For almost 30 days, the chondrocytes retain their spherical shape and produce an aggrecan whose hydrodynamic properties are similar to those of aggrecan molecules in the matrix of articular cartilage, as well as collagen molecules of II, IX and XI types. At the same time, like other suspension cultures, flattened cells are present on the surface of the alginate beads, which produce a small amount of type I collagen molecules that are directly released into the medium and not incorporated into the ECM. In the alginate beads, moderate proliferation of chondrocytes is observed. After 8 months of cultivation in the alginate gel, mature chondrocytes do not lose metabolic activity and continue to synthesize type-II collagen-like collagen and aggrecan.
N. Tanaka and coauthors (1984) investigated the diffusion properties of various natural molecules in the alginate and found that molecules larger than 70 kD do not diffuse through the alginate. Thus, the cultivation of cells in the alginate is suitable for studying the regulation of matrix biosynthesis and the organization of ECM. The availability of cells cultivated in the alginate allows one to investigate the effect of peptide regulatory factors and pharmacological agents on transcriptional, posttranscriptional and translational levels.
Chondrocytes are also cultured in a matrix of collagen fibers I and II types. S. Nehrer and co-authors (1997) compared the functioning of dog chondrocytes in porous collagen-proteoglycan polymeric matrices containing collagens of different types. They found important differences in the morphology of the biosynthetic function of chondrocytes cultured in collagen matrices containing collagen types I and II. Cells in the matrix of collagen type II coiled their spherical shape, while in type I collagen, they had a fibroblast-like morphology. Moreover, in the matrix of type II collagen, chondrocytes produced more glycosaminoglycans. J. Van Susante et al (1995) compared the properties of chondrocytes cultured in the alginate and the collagen (type I) gel. The authors found a significant increase in the number of cells in the collagen gel, but from the 6th day of cultivation, the cells lost a characteristic phenotype, turning into fibroblast-like cells. In the alginate gel, a decrease in the number of cells was observed, but the chondrocytes retained their normal phenotype. In the collagen gel, the number of proteoglycans per cell was significantly higher than in the alginate, however, in the gel, the synthesis of matrix elements was reduced starting from the 6th day of cultivation, while in the alginate the synthesis continued to grow.
A solid three-dimensional fibrin matrix is a natural substance that supports the chondrocytes weighed in it in a differentiated phenotype. The 3D fibrin matrix can also be used as a carrier for chondrocyte transplantation. Advantages of fibrin are the absence of cytotoxicity, the ability to fill the space, the adhesive ability. By histological and biochemical studies, autoradiography, electron microscopy, chondrocytes in the fibrin gel have been found to retain their morphology, multiply and produce a matrix even after 2 weeks of culture. However, G. Homminga and co-authors (1993) reported that after 3 days of culture the disintegration of fibrin begins, the dedifferentiation of chondrocytes progresses.
Suspension of chondrocytes in an artificial (synthetic) ECM
Cartilage implants for reconstructive or orthopedic surgery can be obtained by growing isolated chondrocytes in vitro in a synthetic biocompatible matrix.
Cultured polyglycolic acid chondrocytes proliferate and maintain normal morphology and phenotype within 8 weeks. The chondrocyte-polyglycolic acid complex consists of cells, glycosaminoglycans, collagens, and has an outer collagen capsule. However, in such implants there are two types of collagen molecules - I and II. Implants from dedifferentiated by a series of passages of chondrocytes have a greater number of glycosaminoglycans and collagens than in implants from primarily undifferentiated chondrocytes.
L. Freed and co-authors (1 993b) compared the behavior of human and bull chondrocyte cultures in fibrous polyglycolic acid (HPHC) and in polylactic acid (PPLC). After 6-8 weeks of cultivation of bull chondrocytes in the HSVG or PPLC, the authors observed cell proliferation and cartilage matrix regeneration. In HSBC, the chondrocytes were spherical, located in lacunae surrounded by a cartilaginous matrix. After 8 weeks of in vitro culture, the regenerated tissue contained up to 50% of dry matter (4% of cell mass, 15% of glycosaminoglycans, and 31% of collagen). In PPLK cells were spindle-shaped, a small amount of glycosaminoglycans and collagen. In HSBC, cell growth was 2 times more intense than in PTCA. In in vivo conditions, chondrocytes grown in HPVC and PPLC for 1 to 6 months produced a tissue histologically similar to cartilage. Implants contained glycosaminoglycans, type I and type II collagens.
Fetal bull chondrocytes were cultured in porous high-density hydrophobic and hydrophilic polyethylene. After 7 days of incubation in both substrates, the cells retained a spherical shape, mainly containing type II collagen. After 21 days of cultivation, it turned out that the hydrophilic matrix contains more type II collagen than the hydrophobic matrix.
Cartilage tissue can also be obtained by culturing in a monolayer on Millicell-CM filters. Pre-coating the filters with collagen is necessary for the attachment of chondroits. Histological examination of the culture demonstrates the accumulation of chondrocytes in the ECM containing proteoglycans and type II collagen. Collagen type I in such a culture is not detected. Chondrocytes in the resulting cartilaginous tissue have a spherical shape, but on the surface of the tissue they are somewhat flattened. The thickness of the newly formed tissue increased with time and depended on the initial density of the monolayer of cells. Under optimal culture conditions, the thickness of the cartilaginous tissue reached 110 μm, the organization of its cells and collagen in the surface and deep layers is similar to that of articular cartilage. VKM contains approximately 3 times more collagen and proteoglycans. After 2 weeks of cultivation, the accumulation of the matrix-sa was noted, which made it possible to extract the tissue from the filter and use it for transplantation.
Sims et al. (1996) studied the cultivation of chondrocytes in a polyethylene oxide-gel encapsulated polymer matrix that allows a large number of cells to be transported by injection. Six weeks after injection into the subcutaneous tissue of athymic mice, a new cartilage was formed, which morphologically was characterized by white opalescence similar to hyaline cartilage. The data of histological and biochemical studies indicated the presence of actively proliferating chondrocytes, which produce ECM.
Explantation
Examination of cartilaginous tissue is used to study the processes of ana- and catabolism in it, homeostasis, resorption and repair. Chondrocytes in cartilaginous tissue explants support the normal phenotype and composition of ECM, similar to those in articular cartilage in vivo. After 5 days of cultivation in the presence of serum, a constant level of synthesis and natural degradation is achieved. The tissue resorption can be accelerated in the main culture and culture with the addition of serum by a number of agents, for example, IL-IB, TNF-a, bacterial lipopolysaccharides, retinoic acid derivatives or active oxygen radicals. To study the repair of cartilage, its damage is induced by soluble mediators of inflammation (H 2 O 2, IL-1, TNF-a) or physical rupture of the matrix.
The method of organotypic cultures is a model for studying in vitro effects of isolated external factors on chondrocytes and the surrounding matrix. In vivo, chondrocytes are rarely located in the ECM and do not contact each other. The culture of the explant articular cartilage retains this structural organization, as well as the particular interactions between the chondrocytes and their surrounding extracellular environment. This model is also used to study the effect of mechanical stress, pharmacological agents, growth factors, cytokines, hormones on the metabolism of cartilage.
Another advantage of cartilaginous tissue explantation is the absence of chondrocyte damage by proteolytic enzymes or a mechanical factor, which is inevitable when cells are isolated. Receptors and other membrane proteins and glycoproteins are protected from damaging factors.
Culture of chondrons
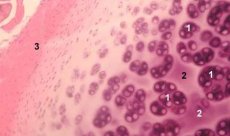
Chondron is a structural, functional and metabolic unit of articular cartilage, consisting of a chondrocyte, its pericellular matrix and a compact filament capsule, and is responsible for the matrix homeostasis. The chondrons are mechanically extracted from the cartilage and collected by several successive low-speed homogenizations. Isolated from zones of different depths of cartilage, chondrons can be divided into four categories: single chondron, coupled chondrons, multiple (three or more) linearly located chondrons (chondron columns), a cluster of chondrons.
Single chondrons are usually found in the middle layers of intact cartilage, paired - on the border of middle and deep layers, linearly located multiple chondrons are typical for deep layers of intact cartilage. Finally, clusters of chondrons consist of randomly organized groups of single and paired chondrons that retain the aggregated state after homogenization. Accumulations of chondrons are large fragments of cartilage, usually containing several chondrons and radially located collagen fibrils, ie, a typical organization characteristic of deep layers of the matrix. Chondrons are immobilized in a transparent agarose, which allows to study their structure, molecular composition and metabolic activity. The chondron - agarose system is considered as a micromodel of cartilage, which differs from the traditional chondrocyte - agarose system in that a natural microenvironment is maintained, there is no need to perform its synthesis and assembly. The culture of chondrons is a model for studying the interactions of cells and matrix in articular cartilage in normal and pathological conditions.
 [22], [23], [24], [25], [26], [27]
[22], [23], [24], [25], [26], [27]
Culture of immortal chondrocytes
To create permanent cell lines, recombinant DNA or oncogen containing viruses are used that can make the cell "immortal". Immortal chondrocytes have the ability to endless proliferation, maintaining a stable phenotype. F. Mallein-Gerin and co-authors (1995) showed that the SV40T-oncogene induces the proliferation of mouse chondrocytes, which continue to stably express collagen II, IX and XI types, as well as articular aggregate and a binding protein. However, such a cell line acquires the ability to synthesize type I collagen when cultured in a monolayer culture or in an agarose gel.
W. Horton and co-authors (1988) described a line of immortal cells with a low level of collagen type II mRNA expression. These cells were obtained by transforming them with a mouse retrovirus containing I-myc- and y-ra-oncogenes. This type of cells is a unique model for studying the interactions of the articular matrix in the absence of type II collagen, as well as the regulation of the synthesis of type II collagen.
The culture of chondropytes with mutated or deleted genes is a convenient model for studying their physiological function. This model is particularly suitable for studying the role of specific molecules in the organization of a cartilaginous matrix or studying the effects of various regulatory factors on the metabolism of cartilage. Chondrocytes with a deleted genome of type IX collagen synthesize collagen fibrils wider than normal, indicating that collagen type IX regulates the diameter of fibrils. As noted in Chapter 1, a mutation of the COLAI gene encoding type II collagen in families with primary generalized osteoarthritis has recently been detected. To study the effect of mutant type II collagen on the articular matrix, R. Dharmrvaram and co-authors (1997) performed a transfection ("contamination" with a foreign nucleic acid) of defective COL 2 AI (arginine at position 519 replaced by cysteine) in human fetal chondrocytes in vitro.
System of cocultures. In the joint, cartilage interacts with cells of other types contained in the synovial membrane, synovial fluid, ligaments, subchondral bone. The metabolism of chondrocytes can be influenced by various soluble factors synthesized by these cells. So, arthritis articular cartilage is destroyed by proteolytic enzymes and free radicals, which are produced by synovial cells. Therefore, models have been developed to study complex interactions between cartilage and surrounding tissues, which are called coculture.
S. Lacombe-Gleise and co-authors (1995) cultivated rabbit chondrocytes and osteoblasts in a co-culture system (COSTAR) in which the cells were separated by a microporous membrane (0.4 μm) allowing exchange between the cells of the two types without any direct contact. This study demonstrated the ability of osteoblasts to stimulate the growth of chondrocytes through soluble mediators.
A.M. Malfait and co-authors (1994) investigated the relationship between monocytes of peripheral blood and chondrocytes. This model is convenient for studying the processes mediated by cytokines, in inflammatory arthropathies (rheumatoid arthritis, seronegative spondylitis, etc.). The authors of the model separated the cells by a protein-binding membrane with pores 0.4 μm in diameter. The study showed that the monocytes stimulated with lipopolysaccharide produced IL-1 and TNF-a, which inhibited the synthesis of aggrecan by chondrocytes and contributed to the degradation of the already synthesized aggrecan aggregates.
K. Tada and co-authors (1994) created a model of coculture in which endothelial cells in a collagen (type I) gel were placed in an inner chamber separated from the outer chamber with a filter with a pore size of 0.4 μm placed in it with chondrocytes. In a state of complete isolation from the outer chamber, human endothelial cells formed tubes in a collagen gel in the presence of EGF or TGF-a. With the simultaneous cultivation of both types of TGF cells, the dependent formation of the tubes by endothelial cells was inhibited. The chondrocyte inhibition of this process was partially eliminated by anti-TGF-beta antibodies. It can be assumed that the TGF-beta produced by the chondrocytes depresses the vascularization of the cartilage itself.
S. Groot and co-authors (1994) simultaneously cultivated chondrocytes from the hypertrophic and proliferative zones of the bone of a 16-day-old fetal mouse with pieces of brain tissue. After 4 days of culture, transdifferentiation of chondrocytes into osteoblasts and the onset of osteoid formation were observed. After 11 days of cultivation, a portion of the cartilage was replaced by bone tissue and the bone matrix was partially calcified. Some neuropeptides and neurotransmitters produced by brain tissue, affect the metabolism of osteoblasts or have receptors on them. Among them, norepinephrine, vasoactive intestinal peptide, peptide associated with the calcitonin gene, substance P and somatostatin can be isolated . Cultured with chondrocytes, pieces of brain tissue can produce some of these factors, which can induce the process of chondrocyte transdifferentiation into osteoblasts.
[28], [29], [30], [31], [32], [33]
The influence of external factors on the culture of chondrocytes
The effect of oxygen tension on the metabolism of chondrocytes
In most cases, chondrocyte cultures develop under conditions of atmospheric oxygen tension. Nevertheless, it is well known that in vivo chondrocytes exist under hypoxic conditions and the oxygen tension varies with different pathological conditions. During the maturation process, significant changes in the blood supply of the epiphyses are observed. Since vascularization varies in different areas of the growth plate, the oxygen tension in them also varies. C. Brighton and R. Heppenstall (1971) demonstrated that in the plate of the tibia in rabbits, the oxygen tension in the hypertrophic zone is less than in the surrounding cartilage. Measurements of some metabolic parameters have shown that chondrocytes are able to react quickly to local changes in oxygen concentration. First of all, with low oxygen tension, its consumption of chondrocytes decreases. With a decrease in oxygen tension from 21 to 0.04%, glucose utilization is increased, glycolysis enzyme activity and lactic acid synthesis are increased. Even with a low oxygen tension, the absolute amount of ATP, ADP, and AMP remains stable. These data indicate the directionality of chondrocyte metabolism to maximize energy conservation. Nevertheless, the synthetic activity, and hence the processes of reparation, change under conditions of hypoxia.
High oxygen tension also affects the metabolism of chondrocytes, causing a decrease in the synthesis of proteoglycans and DNA, degradation of the matrix of cartilage. These effects, as a rule, are accompanied by the production of free oxygen radicals.
Influence of ion concentration and osmotic pressure of the environment on the function of chondrocytes
In native cartilage, the ion concentration differs significantly from that in other tissues: the sodium content in the extracellular medium is 250-350 mmol, and its osmolarity is 350-450 mosmol. When isolating chondrocytes from ECM and incubating them in standard media (DMEM (Dulbecco's Minimal Essential Medium), the osmolarity is 250-280.7 mosmol), the environment surrounding the cell changes dramatically. In addition, the concentration of calcium and potassium in standard media is much lower than in native tissue, and the concentration of anions is much higher.
Addition of sucrose to the medium leads to an increase in its osmolarity and induces a transient intracellular increase in the concentration of H + and calcium anions in the cytosol. Such intracellular changes can influence the processes of chondrocyte differentiation and their metabolic activity. J. Urban et al. (1993) found that the incorporation of 35 8-sulfate and 3 H-proline with isolated chondrocytes incubated in a standard DMEM medium for 2-4 hours was only 10% of that in native tissue. The intensity of synthesis reached a maximum with osmolarity of the extracellular medium of 350-400 mosmol both in the newly isolated chondrocytes and in the explants of the cartilaginous tissue. Moreover, the chondrocyte volume increased by 30-40% after placing isolated cells in a standard DMEM medium of said osmolarity. However, in the cultivation of chondrocytes under nonphysiological osmolarity for 12-16 h, the cells adapt to new conditions, reducing the intensity of biosynthesis in proportion to the osmolarity of the extracellular medium.
P. Borgetti and co-authors (1995) investigated the influence of osmolarity of the extracellular medium on the growth, morphology and biosynthesis of porcine chondrocytes. The authors demonstrated similar biochemical and morphological features of chondrocytes cultured in media with osmolarity of 0.28 and 0.38 mosmol. With osmolarity of 0.48 mosmol during the first 4-6 hours of culture, cell proliferation and protein synthesis were reduced, but subsequently these parameters recovered, which eventually reached control values. When chondrocytes are cultured in a medium with osmolarity of 0.58 mosmol cells lose the ability to maintain the physiological intensity of proliferative processes and after 6 days the amount of chondrocytes significantly decreases. With osmolarity of the medium, 0.58 mosmol, a deep inhibition of protein synthesis is observed. In addition, when cultured in media with osmolarity of 0.28-0.38 mosmol, chondrocytes retain a physiological phenotype, with higher osmolality (0.48-0.58 mosmol), significant changes in the morphology of the cells occur, which is manifested by the loss of a characteristic phenotype, the transformation of chondrocytes into fibroblast-like cells, as well as cell loss, the ability to assemble matrix proteoglycans. The results of this study indicate the ability of chondrocytes to respond to limited osmolality oscillations in the extracellular environment.
The change in the concentration of other ions can also affect the processes of biosynthesis in chondrocytes. Thus, the degree of inclusion of 35 S (sulfate) increases by half with an increase in the concentration of potassium ions from 5 mmol (concentration in a standard DM DM medium) to 10 mmol (concentration in VKM in vivo). Calcium concentration below 0.5 mmol contributed to the production of collagen by mature bull chondrocytes, while a concentration of 1-2 mmol (corresponding to the concentration in the standard DM DM medium) caused a significant decrease in collagen synthesis. A moderate increase in biosynthesis was observed at high levels of calcium (2-10 mmol). Various cations participate in the attachment of chondrocytes to VKM proteins. Thus, magnesium and manganese ions provide attachment to fibronectin and collagen type II, whereas calcium ions do not participate in the attachment of chondrocytes to proteins. Thus, the results of the described studies indicate the influence of changes in extracellular ions of potassium, sodium, calcium and osmolarity of the medium on the biosynthetic function of chondrocytes incubated in standard media.
The influence of mechanical stress on the metabolism of chondrocytes
Immobilization of the joint causes a reversible atrophy of the cartilage, which indicates the need for mechanical stimuli for the normal course of metabolic processes in the ECM. In most cases, the cell culture models used exist under normal atmospheric pressure conditions. M. Wright and co-authors (1996) showed that the mechanical environment affects the metabolism of chondrocytes, the response of cells depends on the intensity and frequency of the compression load. Experiments with loading on explants of intact articular cartilage in vitro demonstrated a decrease in the synthesis of proteins and proteoglycans under the action of a static load, while dynamic loading stimulates these processes. The exact mechanisms for realizing the effect of mechanical stress on the cartilage are complex and are probably related to cell deformation, hydrostatic pressure, osmotic pressure, electrical potential and surface cellular receptors to matrix molecules. To study the effect of each of these parameters, it is necessary to create a system in which one parameter can be independently varied. For example, an explant culture is not suitable for studying cell deformation, but it can be used to study the overall effect of pressure on the metabolic activity of chondrocytes. Compression of cartilage leads to deformation of cells, and is also accompanied by the appearance of a gradient of hydrostatic pressure, electric potential, fluid flow and changes in physicochemical parameters such as water content in the matrix, electrical charge density, and osmotic pressure level. Cell deformation can be studied using isolated chondrocytes immersed in an agarose or collagen gel.
Several systems have been developed to study the effect of mechanical stimulation on the culture of chondrocytes. Some researchers use systems for this purpose in which the pressure is applied to the cell culture through the gaseous phase. Thus, JP Veldhuijzen et al. (1979), using a pressure above atmospheric at 13 kPa at a low frequency (0.3 Hz) for 15 min, observed an increase in the synthesis of cAMP and proteoglycans and a decrease in DNA synthesis. R. Smith and co-authors (1996) showed that an intermittent exposure of a primary chondrocyte culture to a hydrostatic pressure (10 MPa) at a frequency of 1 Hz for 4 hours caused an increase in the synthesis of aggrecan and collagen type II, while constant pressure did not affect these processes. Using a similar system, M. Wright and co-authors (1996) reported that cyclic pressure on cell culture is associated with hyperpolarization of the cell membrane of chondrocytes and activation of Ca 2+ -dependent potassium channels. Thus, the effects of cyclic pressure are mediated by ion channels, activated by stretching, in the chondrocyte membrane. The response of chondrocytes to hydrostatic pressure depends on the conditions of cell culture and the frequency of the applied load. Thus, the cyclic hydrostatic pressure (5 MPa) reduces the inclusion of sulfate in the monolayer of chondrocytes at a frequency of 0.05, 0.25 and 0.5 Hz, while at a frequency of more than 0.5 Hz, the inclusion of sulfate in the explant cartilage increases.
M. Bushmann et al. (1992) reported that chondrocytes in an agarose gel alter biosynthesis in response to static and dynamic mechanical stress in the same way as the cultured intact organ. The authors found that the mechanical load generates a hyperosmotic stimulus followed by a decrease in pH in the chondrocytes.
The effect of mechanical stretching can be studied on a culture of cells immersed in a gel. The stretching force can be created using a computer-controlled vacuum. When the system is in a vacuum of a certain degree, the bottom of the Petri dish with the cell culture is extended by a certain amount, the deformation is maximal at the edges of the bottom of the cup and is minimal in the center. Stretching is transmitted and cultured in a petri dish of chondrocytes. With the help of this method, Holm-vall and co-authors (1995) have shown that expression of mRNA of α 2 -integrin is increased in cultured collagen (type II) gel chondrosarcoma cells . And 2 p g integrin is able to bind to type II collagen. It is considered as a mechanoreceptor, since it interacts with actin-binding proteins, thereby connecting the ECM and the cytoskeleton.
Effect of pH on Chondrocyte Metabolism
The pH of the interstitial fluid of ECM of the cartilaginous tissue is more acidic than in other tissues. A. Maroudas (1980) determined the pH of the articular cartilage at 6.9. W. Diamant and co-authors (1966) found a pH of 5.5 in pathological conditions. It is known that chondrocytes live at low PO2, which indicates the important role of glycolysis (95% of total glucose metabolism) in the metabolism of these cells; glycolysis is accompanied by the production of a large amount of lactic acid.
In addition to acidification of the environment by the products of glycolysis, the matrix components themselves are of great importance. A large amount of fixed negative charge on proteoglycans modifies the extracellular ionic composition: a high concentration of free cations (for example, H +, Na +, K + ) and a low concentration of anions (for example, O2, HCO3) are noted. In addition, under the influence of a mechanical load, water is expelled from the ECM, which leads to an increase in the concentration of fixed negative charges and the attraction of more cations to the matrix. This is accompanied by a decrease in the pH of the extracellular medium, which affects the intracellular pH, thereby modifying the metabolism of chondrocytes. R. Wilkin and A. Hall (1995) studied the effect of the pH of the extracellular and intracellular medium on the biosynthesis of the matrix by isolated bull chondrocytes. They observed a dual modification of matrix synthesis with a decrease in pH. A slight decrease in pH (7.4
[34], [35], [36], [37], [38], [39], [40]
Effect of the composition of the medium for cultivation on the metabolism of chondrocytes
The medium for culturing the chondrocytes must correspond to the experimental conditions. In recent years, calf serum has been used to optimize the culture conditions. However, when using serum, a number of important points should be considered:
- external growth of cells from the periphery of tissue in organ cultures,
- the variability of the composition of sera of various series,
- presence of unknown components in them,
- increased risk of interference, artifacts in the study of the influence of various biological factors on the metabolic activity of cells.
An example of the latter is the study of the effect of EGF on cartilage chondrocytes in rats. EGF stimulated the incorporation of 3 H-thymidine and an increase in DNA content in the culture. This effect was more pronounced at low serum concentrations (<1%), but at high concentrations (> 7.5%) the effect disappeared.
It is well known that levels of synthesis and degradation in DMEM enriched with calf serum are significantly increased compared to in vivo conditions. Differences between in vivo and in vitro metabolism can be caused by differences between the synovial fluid and the environment in which the cells are cultured. D. Lee et al. (1997) cultivated chondrocytes of young bulls in agarose using a nutrient medium containing DMEM enriched with 20% calf serum and a large amount of normal allogeneic synovial fluid. The presence of synovial fluid in the medium induced an increase in the number of proteoglycans, up to 80% of the total amount of synovial fluid. The results obtained indicate that synovial fluid in culture induces a metabolic rate similar to that in vivo, with a high level of synthesis of glycosaminoglycans and a low level of cell division.
G. Verbruggen and co-authors (1995) showed that the synthesis of 35 S-arrpeKaHa by human chondrocytes cultured in agarose in DMEM without serum was 20-30% of the level of synthesis observed in DMEM enriched with 10% calf serum. The authors determined the extent to which IGF-1, IGF-2, TGF-P or insulin restore the production of aggrecan in serum-free media. The authors concluded that 100 ng / ml of insulin, IGF-1 or IGF-2 partially reduced the synthesis of aggrecan to 39-53% of the control level. With a combination of these factors, no synergistic or cumulative phenomena have been identified. At the same time, 10 ng / ml of TGF-P in the presence of 100 ng / ml of insulin stimulated the synthesis of aggrecan up to 90% or more from the reference level. Finally, serum transferrin, alone or in combination with insulin, did not affect the synthesis of aggrecan. When calf serum was replaced with bovine serum albumin, the aggregate content of aggrecan was significantly reduced. Enrichment of the medium for insulin culture, IGF or TGF-P partially restored the cells' ability to produce aggrecan aggregates. In this case, IGF-1 and insulin are able to maintain homeostasis in cell cultures. After 40 days of cultivation in medium enriched with 10-20 ng / ml IGF-1, the synthesis of proteoglycans was maintained at the same level or even higher than in the medium containing 20% calf serum. Catabolic processes proceeded more slowly in a medium enriched with IGF-1 than in a medium enriched with 0.1% albumin solution, but somewhat faster in a medium enriched with 20% serum. In long-living cultures, 20 ng / ml IGF-1 maintains a stable state of cells.
D. Lee and co-authors (1993) compared the effect of the culture medium composition (DMEM, DMEM + 20% calf serum, DMEM + 20 ng / ml IGF-1) on DNA synthesis in a cartilage explant culture, monolayer culture and in agarose suspension . When cultured in agarose in the presence of serum, the authors observed a tendency to group chondrocytes into large clusters. Cells cultured without serum or with IGF-1 were stored in a round-shaped agarose, assembled into small groups, but did not form large aggregates. In the monolayer, the synthesis of DNA was significantly higher in serum-containing media than in the medium enriched with IGF-1; The synthesis of DNA in the latter was much higher than in the unenriched environment. In the cultivation of chondrocytes in a suspension in agarose in an unenriched medium and in media with IGF-1, no differences in DNA synthesis were observed. At the same time, culturing the chondrocyte suspension in agarose in a medium enriched with serum was accompanied by an increased incorporation of the radionucleotide 3 H-thymidine as compared to other media.
Vitamin C is necessary for the activation of enzymes involved in the formation of a stable spiral structure of collagen fibrils. Chondrocytes, deficient with respect to ascorbic acid, synthesize under-hydroxylated non-helical precursors of collagen, which are slowly secreted. The introduction of ascorbic acid (50 μg / ml) causes hydroxylation of collagen types II and IX and their secretion in normal amounts. The addition of vitamin C did not affect the level of synthesis of proteoglycans. Consequently, the secretion of collagen is regulated independently of the secretion of proteoglycans.